
放射齒目
| 放射齒目 | |
|---|---|

| |
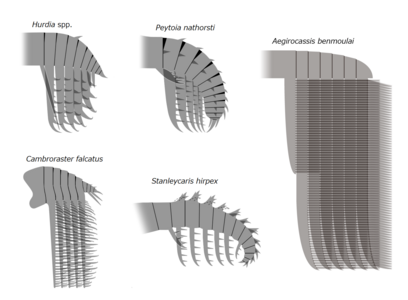
| 七個放射齒目物種的復原圖(未依比例顯示) 由左至右,自上而下分別為: | |
| 科學分類 | |
| 界: | 動物界 Animalia |
| 門: | 節肢動物門 Arthropoda |
| 綱: | †恐蝦綱 Dinocaridida |
| 目: | †放射齒目 Radiodonta Collins, 1996 |
| 科 | |
|
不屬於任何科的屬 | |
| 寒武紀大爆發 |
|---|
放射齒目(學名:Radiodonta),又稱射口類[1]、奇蝦類[1]、奇蝦動物[2],是節肢動物一個已滅絕的基群(有時亦被劃入葉足動物門下),屬於恐蝦綱,其下共計19個有效屬。放射齒目的化石在歐、亞、非、北美等大洲,以及澳洲等地皆有發現,涵蓋的時間跨度由寒武紀一直延伸至泥盆紀,長達1.2億年;最著名的放射齒類動物奇蝦,便是寒武紀的代表性生物之一。
放射齒目體型從不及10公分至2公尺皆有,身型可分為頭、軀幹兩個體節。其中,頭部長有複眼和骨化的前附肢,以及齒環繞四周的口錐;軀幹則分為許多小節,每節各有一對由肌肉束控制的扇鰭。此類群也有由腸道、消化腺組成的消化系統,和眼節附近的大腦等結構組成的神經系統。據其形態推測,放射齒目可以利用扇鰭在水中游動,有的甚至可以利用其尾扇快速轉向。覓食時,它們有的能利用前附肢抓取獵物,有的能從海底泥中濾取食物碎屑,還有的能夠濾食浮游生物。
放射齒目作為研究節肢動物起源和早期系統發育的重要類群,其分類方式在分類學研究中獲高度關注。然而,它們的化石極難保存下來,因此出現過不少分類錯誤。[3]
詞源與俗名
[編輯]放射齒目的學名「Radiodonta 」來自拉丁語「radius」(輪輻)和希臘語「odoús」(齒),這指的是其放射狀分布的牙齒[4],但並非所有該目生物均有此特徵[5][6]。
本目的俗名包含「radiodonts」[5][7][8]、「radiodontans」[6][9]、「radiodontids」[4]。此外,鑑於放射齒目至今19個有效屬中絕大多數都屬於奇蝦亞目(Anomalocarida),一些英文文獻也會直接稱之為「anomalocaridids」[10][11][12] 或 「anomalocarids」[13],中文又譯為奇蝦類[1]或奇蝦動物[2]。
定義
[編輯]1996年,加拿大古生物學家戴斯蒙·柯林斯建立恐蝦綱與放射齒目,當時柯林斯將放射齒目定義為:[4]
放射齒目是兩側對稱、體型修長的節肢動物,其未礦化的表皮在口顎、爪子處尤為堅韌。放射齒目的身體可分為兩個體段,類似螯肢亞門節肢動物的頭胸部和腹部。通常來講,前體段沒有明顯的外部分節,口前長有一對爪,一對突出的複眼,以及位於腹側帶有輻射狀牙齒。有幾種放射齒目動物還擁有額外的一層牙齒,以及三、四對後口咀嚼式附肢。軀幹分節,通常為13節,節的兩側生有片狀的構造用以游泳,以及用以呼吸的鰓。尾部可能有三片向外突出的構造。有些物種軀體上生有咀嚼式附肢。
2014 年,學者以系統發生學的角度定義了放射齒目:任何親緣關係比堪察加擬石蟹更接近加拿大奇蝦皆為放射齒目[13]。2019 年,學者以形態學的角度定義了放射齒目:頭甲複合結構(head carapace complex) 包含 H骨片(H-element) 和 P骨片(P-element);前附肢棘(endites) 生有前附肢輔棘(auxiliary spines);前端的扇鰭(flaps) 或剛毛葉(setal blades) 退化;軀體梯形,前端較高,往後漸低[8]。

特徵
[編輯]
絕大多數的放射齒目動物都比其他的寒武紀動物大得多,身長通常介於30至50厘米之間[7]。已知最大的放射齒目是奧陶紀的本莫拉海神盔蝦(Aegirocassis benmoulai),其最大體長可達兩公尺[12]。最小的放射齒目之一是8厘米的里拉琴蟲屬,有爪里拉琴蟲(L. unguispinus) 的幼體只有1.8公分[14]。
放射齒目動物的身體可以分成兩個部分:頭部和軀幹。頭部只有一節體節[15],稱作「眼節」(ocular somite),眼節被骨片覆蓋,覆蓋眼節的骨片統稱「頭甲複合結構」(head carapace complex),眼節前側生有分節的附肢,腹側生有口器(口錐,oral cone),還有眼柄和複眼。軀幹呈梯形,越往後越低,每節體節都生有一對扇鰭(flap) 和名為剛毛葉(setal blades) 的鰓狀結構[8]。
前附肢
[編輯]



頭前側生有一對前附肢(frontal appendage),前人研究中曾稱該對附肢為「爪」(claws)、「取食足」(feeding appendages)、「大附肢」(great appendage),但近期的文獻不再使用大附肢一詞,因為學者認為放射齒目的前附肢與大附肢綱的大附肢不同源[15]。前附肢骨化且分節,大部分附肢上有一節一節的稱作分節(podomeres) 腹側上有棘,稱作「前附肢棘」(endites),該棘的前側和後側可能會額外生有數排棘,稱作「前附肢輔棘」(auxiliary spines)[16]。前附肢可分為兩部分,梗節(shaft) 和端節(distal articulated region)[16],梗節為靠近頭部的部分,端節為延伸出去的部分。有些物種腹部具有三角狀的節間膜,可使軀體更為柔軟靈活[17]。放射齒目眼前節(與第一大腦相關)與有爪動物門的主觸角和真節肢動物(Euarthropoda) 的上唇同源,都起源於眼節;而上述結構則與螯肢亞門的螯肢和其他節肢動物的觸角和大附肢不同源,這些結構起源於第一眼後節(與第二大腦相關)[11][15]。前附肢的形態因物種而異,尤其前附肢棘差異更大,因此前附肢和前附肢棘為分辨物種的重要特徵[16]。甚至有些放射齒目的物種是單靠前附肢標本發表的,例如高蹺雙棘蝦(Ursulinacaris grallae)、抱怪蟲屬中的史蒂芬山抱怪蟲(Amplectobelua stephenensis)[16][17]。
-
 奇蝦科、抱怪蟲科和其他近緣物種的前附肢
奇蝦科、抱怪蟲科和其他近緣物種的前附肢 -
 篩蝦科的前附肢
篩蝦科的前附肢 -
 赫德蝦科的前附肢
赫德蝦科的前附肢



口器
[編輯]

口器位於頭部的腹側,前附肢的著生點之後。口器周圍環繞著一圈齒,這種形式的口器稱作「口錐」(oral cone),舊文獻中稱作「顎(jaws)」。有些物種會有三或四齒增大,讓口器外形呈現三向(如奇蝦屬)或四向(如赫德蝦屬)的輻射對稱[14][18]。齒的內部朝向口器開口的地方生有刺,有些赫德蝦科的齒內刺比其他物種來得多層[8][10]。抱怪蟲科的口器細節復原圖是推測而來,實際的口器可能為非典型的輻射對稱[5][6]。
頭部骨片、複眼和軀體
[編輯]

頭甲複合結構由三個骨片組成:中央的 H 骨片(前側骨片或頭盾)和一對 P 骨片(兩側骨片),H 骨片包覆頭部背側,P 骨片包覆頭部兩側的腹面[8]。有些物種的兩個 P 骨片和 H 骨片相連,形成一個狹窄的前側延伸,稱作「P 骨片頸」(P-elements neck)或「喙橋」(beak)[8][10]。奇蝦科和抱怪蟲科的頭甲複合結構較小,且呈卵形[8][6],赫德蝦科的較大[8]。頭部生有一對眼柄,著生於 H 骨片和 P 骨片後側的間隙[8][10],眼柄末端生有複眼,學者推測眼柄可移動[19]。
不同於原始描述的是,放射齒目的體軀分節十分明顯[8][9][12],沒有任何放射齒目的具有足狀附肢[20](除了瓜肢蟲屬,但該屬是否屬於放射齒目還未有定論[12][21])。體軀分很多節,呈梯形,由前向後漸低,前三或前四節隘縮,形成頸部[8]。



軀體兩側生有鰭狀附肢,稱作「扇鰭」(flaps),有些文獻稱作「側鰭」(lateral flaps)和「葉」(lobes),通常一個體節會有一對扇鰭,扇鰭的前側會和前一個扇鰭的後側稍微重疊,有些赫德蝦科的物種背側會生有一列不和前一個扇鰭重疊的小扇鰭[12]。扇鰭生有無數條線狀結構,稱作「線狀支撐結構」(strengthening rays[8][9][12])。頸上的扇鰭稱作「前鰭」(anterior flaps)或「頸鰭」(neck flaps),前扇鰭明顯退化。有些物種具有「咀嚼式結構」(gnathobase-like structures, GLSs),咀嚼式結構著生於前扇鰭的基部[5][6]。背上有列狀的「剛毛葉」(setal blades or lamellae)[8],剛毛葉是由無數的細長結構排列而成,似鰓,剛毛葉附蓋在體節的背側[12]。
腹側的扇鰭可能與真節肢動物的內足(endopod) 和葉足動物的葉足(lobopod) 同源,而背側的扇鰭和剛毛葉可能與真節肢動物的外足(exopod)和葉足動物背側生有鰓的扇鰭(gill-bearing dorsal flaps)同源[12][22]。軀體末端的構造因物種而異,可能會有以下形態:一至三對的尾扇[8][20]、兩條細長的尾毛(furcae)[8] [23]、一條延長的尾部結構(terminal structure[20]) 或沒有其他突出物的圓滑尾端[12]。
-
 赫德蝦科的三個成員:海神盔蝦(上)、皮托蝦(左下)與赫德蝦(右下),三者皆生有背部的翼鰭和發達的頭甲複合結構。
赫德蝦科的三個成員:海神盔蝦(上)、皮托蝦(左下)與赫德蝦(右下),三者皆生有背部的翼鰭和發達的頭甲複合結構。 -
 加拿大奇蝦(Anomalocaris canadensis),其頭甲複合結構呈現卵形,體末生有三對尾鰭和一條尾鬚(terminal tailpiece)。
加拿大奇蝦(Anomalocaris canadensis),其頭甲複合結構呈現卵形,體末生有三對尾鰭和一條尾鬚(terminal tailpiece)。 -
 雙肢抱怪蟲(Amplectobelua symbrachiata),具有咀嚼式結構和兩條尾須。
雙肢抱怪蟲(Amplectobelua symbrachiata),具有咀嚼式結構和兩條尾須。



內部構造
[編輯]
有些放射齒目化石可見肌肉束、消化系統和神經系統。每個體節的兩側生有一束發達的肌肉,連結到腹側的扇鰭[11][20]。前腸和後腸比中腸來的大,中腸狹窄且生有六對憩室(消化腺)[9][20][24]。真節肢動物的大腦分成三部分,有爪動物的大腦分成二部分,而放射齒目的大腦只有一個部分,位於眼節。前附肢的神經著生於大腦的前側,複眼的神經著生於大腦的兩側[11][15]。大腦的後側有一對未癒合的神經節,通過頸部向軀體延伸[11]。
生存年代
[編輯]放射齒目的化石主要產地在中國、美國、加拿大、波蘭、澳大利亞、摩洛哥等地的寒武紀地層,其年代與最早的真節肢動物化石年代,都晚於目前已知最古老的三葉蟲化石,暗示了其間存在化石證據斷層[25]。
-

-

-
 早寒武皮托蝦的前附肢化石保存樣貌
早寒武皮托蝦的前附肢化石保存樣貌

1975年,卡齊米拉·倫茲翁(Kazimiera Lendzion)發表了一塊來自波蘭中東部的扎維申地層(Zawiszyn Formation)的化石,化石年代屬於較早的寒武紀第三期,比澄江化石地的年代還早。當時此化石被誤認為三葉蟲,1977年才確定是放射齒目的物種,認定為早寒武皮托蝦。2009年,學者們發表在早泥盆世(布拉格期至埃姆斯期)的洪斯呂克板岩生物群中發現了巴氏大鰭盔蝦,並將其劃入放射齒目,將已知放射齒目的生存年代往後延續了約1億年[26]。2011年5月,人們在摩洛哥奧陶紀地層發現了新的放射齒目化石[27]。2015年,這批化石被證實屬放射齒目並命名為「班莫拉海神盔蝦」。
生態學
[編輯]生理學
[編輯]放射齒目被認為是在海底生活的生物,且具有游泳能力,從形態上來看,放射齒目可能經常游動。根據肌肉發達且彼此重疊的腹部扇鰭推測,放射齒目可以透過扇鰭游泳,其運動方式可能呈波浪狀,類似現生魟魚和墨魚[28]。背側的扇鰭或有些物種中的尾扇則可以改變行進的方向,或在游泳時穩定自身[12][26]。根據奇蝦屬的尾扇推測,奇蝦可以在水中快速轉向[29]。剛毛葉帶有皺褶的針狀結構,可以擴大面積,學者推測其可能是鰓,用來交換氧氣以供呼吸[12][20]。
有時候脫落骨化結構(如前附肢和頭甲複合結構)會大量聚集,學者推測放射齒目可能會群聚在一起蛻皮[8][12],其他寒武紀的節肢動物(如三葉蟲)也有群聚蛻皮的行為[30]。
食性
[編輯]
放射齒目的物種具有多樣化的覓食策略,包括掠食、篩食底泥或濾食[7][31]。掠食者包括奇蝦與抱怪蟲,會利用前附肢去攫取獵物,抱怪蟲的前附肢甚至長有粗壯的內葉可以像螯那樣鉗住獵物[6][17][21],較小的頭甲及較大的關節膜讓這些掠食性放射齒目物種的前附肢更為靈活[23]。底泥篩食者如赫德蝦與皮托蝦具有較厚實的前附肢,附肢內葉為鋸齒狀並向身體中軸彎曲,形成類似篩子的構造可以將底泥中的食物碎屑濾出並推向口錐[8],其他放射齒目則有不同針對食底泥的演化適應,例如顯爪蝦屬就具有圓頂狀的 H 骨片,外型類似鱟的頭胸甲[8]。濾食者如篩蝦屬與海神盔蝦屬(Aegirocassis) 其前附肢的內葉具有密集排列的柔韌輔助棘,可以過濾出體型最小僅 0.5 mm 的浮游動物及浮游植物[13][12]。胡桃鉗蟲屬(Caryosyntrips) 具有和其他放射齒目相異的前附肢,其內葉為向內相對,類似於剪刀,可用於攫取或切斷獵物[17][32]。



根據口器形態,學者推測放射齒目可能以吸吮或啃咬的方式進食,或兩者並行[8][18][31],和前附肢一樣,不同形態的口器也暗示著放射齒目的食性多樣化。如奇蝦屬的口器為三向輻射對稱,齒不規則且凹凸不均,口器開口較小,這些特徵暗示著奇蝦屬以小型且活動力強的獵物為食[31]。赫德蝦屬和顯爪蝦屬的口器開口較大,且齒數較多,暗示著牠們可能以較大的食物為食[8][31]。
分類
[編輯]系統發生學
[編輯]
| ||||||||||||||||||||||||||||||||||||||||||||||||
| 放射齒目和其他蛻皮動物分類的系統發生樹[33] |




多數分析顯示,放射齒目是節肢動物門的基群[7][8][13][10][11][12],且為次足類(Deuteropoda) 的姊妹群,次足類包含一些較後期的基群(如撫仙湖蟲目、金臂蟲目)和真節肢動物(如類肢綱、螯肢亞門和有顎亞門[33])。許多放射齒目生物上類似節肢動物的特徵支持了以上假說,這些特徵包含複眼[19]、消化腺[24]、前附肢的分節[8][33]、軀體背側和腹側的附肢(後來演化成節肢動物的足[8][12])、被外表皮覆蓋的前腸及後腸[20],和第一大腦前側的骨片[34]。隘縮的頸部和咀嚼式結構後來可能演化成節肢動物的頭部,其複雜的頭部是由數個體節癒合而來[6][15]。


放射齒目和真節肢動物的基群為厭惡蟲屬(Pambdelurion)、宣揚爪蟲屬(Kerygmachela)和歐巴賓海蠍屬,以上三屬是一類形似放射齒目的恐蝦綱生物,常被稱作「帶鰓的葉足動物[12][33]」。帶鰓的葉足動物具有扇鰭、消化腺和如同放射齒目一般特化的前附肢,但帶鰓的葉足動物的前附肢沒有分節,且其扇鰭腹側生有葉足[12]。歐巴賓海蠍屬具有類似放射齒目的眼柄和複眼、尾扇、剛毛葉,和類似真節肢動物位於後側的嘴[33]。比帶鰓的葉足動物更基群的是大網蟲屬(Megadictyon) 和尖山葉足蟲屬(Jianshanopodia)[33], 以上兩屬是葉足動物,生有發達的前附肢和消化道,但沒有扇鰭,其形態介於「葉足動物」與「放射齒目和真節肢動物」之間,暗示著全部的節肢動物可能由併系的葉足動物演化而來,全部的節肢動物包含泛節肢動物的緩步動物門和有爪動物門[15]。
舊時研究不認為放射齒目是節肢動物的基群,而認為其為透過趨同演化而來的環神經動物(此假說乃是根據放射齒目與環神經動物相似的環狀口器[35]);經比較放射齒目的前附肢、大附肢綱的大附肢和螯肢亞門的螯肢後,學者認為螯肢亞門與大附肢綱為姊妹群[36];曾有學者認為具有接近真節肢動物特徵的巴氏大鰭盔蝦(Schinderhannes bartelsi)屬於赫德蝦科[7][8][13][12][33],且是最接近真節肢動物的放射齒目[26],但後來的研究既不支持該種為赫德蝦科,也不支持該種最接近真節肢動物。環狀排列的口器並非環神經動物特有的,而是經過趨同演化而來,也可能是或為蛻皮動物的祖徵(因其同時也在泛節肢動物的緩步動物門與部分有爪動物門中出現[37])。學者認為大附肢綱的大附肢和第二大腦有關[38][39],因此和放射齒目中和第一大腦有關的前附肢非同源[11][15]。僅在一具大鰭盔蝦化石上所發現的真節肢動物特徵有待商榷,其化石可能具有其他類似放射齒目的特徵[33]。


內部分類
[編輯]以前,放射齒目包括恐蝦綱下所有的種類,放射齒目中的所有種類又歸到一個科裡——奇蝦科[10][21],因此即使後來該類群重新分類了,「奇蝦」一詞仍偶爾作為俗稱用來指代整個放射齒目[11][12]。基群的恐蝦科厭惡蟲屬、宣揚爪蟲屬和歐巴賓海蠍屬最近從放射齒目中被移除[7][8][13][12],大多數的放射齒目物種被重新分為三個科:抱怪蟲科、篩蝦科(Tamisiocarididae)[7][8],和赫德蝦科。加上奇蝦科,這四個現行的放射齒目的科可能組成了演化支奇蝦亞目[13]。
放射齒目最初描述時包括奇蝦屬、Laggania(後來改稱皮托蝦屬)、赫德蝦屬、Proboscicaris、抱怪蟲屬、瓜肢蟲屬和似皮托蝦屬[4]。但現在,Proboscicaris被視為赫德蝦屬的同物異名,似皮托蝦屬則被重新分入大附肢綱[10][12][20]。瓜肢蟲屬在放射齒目中的位置並不明確,因為該屬未被拿來進行支序發生學研究[8],其與放射齒目和真節肢動物的支序發生樹呈多分歧狀,且還未被釐清[12]。


如果將瓜肢蟲屬包含在放射齒目中,瓜肢蟲屬與胡桃鉗蟲屬是放射齒目中最基群的類群。不過這兩個類群現在一般不認為屬於奇蝦類。抱怪蟲科內的關係尚未釐清,常有學者質疑里拉蟲屬和蘭氏蝦屬是否屬於抱怪蟲科[5][8]。赫德蝦科的多樣性較高,多個共演徵支持其為單系群的假說,其共演徵包含前附肢的端節共有 5 節,且前附肢棘形態幾乎相同[8][16],很多研究認為篩蝦科為赫德蝦科的姊妹群[7][8][13][12]。
- †放射齒目 Radiodonta
- †奇蝦亞目 Anomalocarida
- †似奇蝦屬 Paranomalocaris[40](一些研究將其歸入奇蝦科[23])
- †薄刃蝦屬 Laminacaris(一些研究將其歸入抱怪蟲科[7])
- †奇美拉薄刃蝦 L. chimera
- 開拓蝦屬 Innovatiocaris[42]
- †帽天山開拓蝦 I.maotianshanensis
- †多刺開拓蝦 I.? multispiniformis
- †奇蝦科 Anomalocarididae
- †抱怪蟲科 Amplectobeluidae
- †篩蝦科 Tamisiocarididae(舊稱鯨蝦科 Cetiocaridae)
- †赫德蝦科 Hurdiidae
- †海神盔蝦屬 Aegirocassis
- †本莫拉海神盔蝦 A. benmoulai
- †皮托蝦屬 Peytoia
- †納氏皮托蝦 P. nathorsti
- †早寒武皮托蝦 P. infercambriensis
- †大鰭盔蝦屬 Schinderhannes
- †巴氏大鰭盔蝦 S. bartelsi
- †斯坦利蝦屬 Stanleycaris
- †耙肢斯坦利蝦 S. hirpex
- †清江斯坦利蝦 S. qingjiangensis[43]
- †赫德蝦屬 Hurdia
- †維多利亞赫德蝦 H. victoria
- †三角赫德蝦 H. triangulata
- †巨盔蝦屬 Titanokorys
- †蓋氏巨盔蝦 T. gainesi
- †小熊蝦屬 Ursulinacaris
- †高棘小熊蝦 U. grallae
- †帕凡特蝦屬 Pahvantia
- †矛刺帕凡特蝦 P. hastata
- †偽窄齒蝦屬 Pseudoangustidontus
- †雙排刺偽窄齒蝦 P.duplospineus
- †篩濾偽窄齒蝦 P. izdigua
- †寒武耙蝦屬 Cambroraster
- †鐮刺寒武耙蝦 C. falcatus
- †心蝦屬 Cordaticaris
- †線紋心蝦 C. striatus
- †口刺蝦屬 Buccaspinea[44]
- †庫氏口刺蝦 B. cooperi
- †海神盔蝦屬 Aegirocassis
- †奇蝦亞目 Anomalocarida
Vinther 等人在 2014 年對放射齒目進行了深入的譜系學分析,同年稍晚叢培允等人添加了有爪里拉琴蟲對研究進行了擴充[11]。 Van Roy 等人於 2015 年對此進一步研究,並加入了優美瓜狀肢蟲(Cucumericrus decoratus) 和本莫拉海神盔蝦[12]。
|
†奇蝦科 Anomalocarididae |

不再屬於放射齒目的類群
[編輯]
由於發現的化石樣本稀少,有些生物曾被錯誤地歸入放射齒目之下:
- 人們目前只發現了全牙蟲的一些牙齒化石。1994年,這些化石被認為是放射齒目動物的口錐[52],到了2006年時人們認定其為一個新屬,將其歸為鰓曳動物[51]。雅各·溫瑟爾(Jakob Vinther)等人於2016年的一份研究中指出,厭惡蟲的口器與全牙蟲極為相似[53]。
發現史
[編輯]放射齒目的化石在歐、亞、非、北美等大洲,以及澳洲等地皆有發現[58]。
大多放射齒目的表皮柔軟,由於身體只有部分骨化,屍體和蛻殼很容易散掉[59]。因此完整的全身化石十分罕見,研究者有時會把不完整的放射齒目化石誤認作其他生物,或把多種放射齒目誤認為一種。通常,研究人員只能找到前附肢、口器、甲殼、扇鰭等,其中前附肢的化石能提供大量有用信息,有不少類群的建立是基於前附肢化石的發現之上[60][4]。
-
 加拿大奇蝦的前附肢化石。以前常被認作葉蝦類的腹部。
加拿大奇蝦的前附肢化石。以前常被認作葉蝦類的腹部。 -
 同屬放射齒目的動物的近乎完整化石(包含前附肢)。
同屬放射齒目的動物的近乎完整化石(包含前附肢)。

加拿大奇蝦和納氏皮托蝦是這方面的典型案例,二者最早發掘出來的都只有不完整的局部化石。其中,前者起初只是發現了前附肢,但研究人員當時誤認為是某種蝦子的腹部,也因此給它取名為「奇異的蝦子」[24]。目前所發現的加拿大奇蝦化石都只有腹部,頭部的化石則至今仍無下落。後者的口器化石則被誤認為是一種水母,並被命名為納氏皮托蝦(Peytoia nathorsti)。而納氏皮托蝦的胴體被誤認為是一種海綿或是海參,被命名為 Laggania cambria[61]。最初這些化石並不認為互相有任何關聯,一直到了1980年代挖掘到了完整的放射齒目物種化石,才理解到原本化石復原上的錯誤。在這之後科學家陸續發表了數個放射齒目的屬與種,將前附肢的細節、尾鰭的有無、口器牙齒的配置等物種差異區別開來[62]。此外,由於大多化石的發現並不完整,部分被認為屬於某物種的化石之後被發現其實屬於其他物種,著名的例子包括有些奇蝦的牙齒化石,在之後被發現其實是皮托蝦的牙齒[63]、以及赫德蝦的頭部骨片化石被發現其實屬於帕凡特蝦[5]。
依照慣例,學名必須使用並沿用第一次公布的名稱,因此即使「奇異的蝦子(Anomalocaris)」最早是針對前附肢化石錯誤復原的命名,但仍然沿用至今作為奇蝦的正式學名。而依據口器牙齒化石命名的模式種皮托蝦(Peytoia)被視為是模式種,而 Laggania 則被視為是異名[63]。
參考資料
[編輯]- ^ 1.0 1.1 1.2 朱茂炎, 趙方臣, 殷宗軍, 曾晗, 李國祥. 中国的寒武纪大爆发研究: 进展与展望. 中國科學: 地球科學. 2019, 49 (10): 1455–1490 [2020-05-02]. ISSN 1674-7240. doi:10.1360/SSTe-2019-0183. (原始內容存檔於2020-01-07).
- ^ 2.0 2.1 叢培允, 侯先光. 澄江动物群奇虾动物的分异度. 中国古生物学会第十次全国会员代表大会暨第25届学术年会——纪念中国古生物学会成立80周年论文摘要集. 中國古生物學會. 2009.
- ^ Pates, Stephen; Daley, Allison C.; Butterfield, Nicholas J. First report of paired ventral endites in a hurdiid radiodont. dx.doi.org. 2019-06-12 [2023-10-09].
- ^ 4.0 4.1 4.2 4.3 4.4 Collins, Desmond. The "evolution" of Anomalocaris and its classification in the arthropod class Dinocarida (nov.) and order Radiodonta (nov.). Journal of Paleontology. 1996, 70 (2): 280–293. doi:10.1017/S0022336000023362.
- ^ 5.0 5.1 5.2 5.3 5.4 5.5 5.6 Cong, Pei-Yun; Edgecombe, Gregory D.; Daley, Allison C.; Guo, Jin; Pates, Stephen; Hou, Xian-Guang. New radiodonts with gnathobase-like structures from the Cambrian Chengjiang biota and implications for the systematics of Radiodonta. Papers in Palaeontology. 2018, 4 (4): 605–621. ISSN 2056-2802. doi:10.1002/spp2.1219 (英語).
- ^ 6.0 6.1 6.2 6.3 6.4 6.5 6.6 Cong, Peiyun; Daley, Allison C.; Edgecombe, Gregory D.; Hou, Xianguang. The functional head of the Cambrian radiodontan (stem-group Euarthropoda) Amplectobelua symbrachiata. BMC Evolutionary Biology. 2017, 17 (1): 208. ISSN 1471-2148. PMC 5577670
 . PMID 28854872. doi:10.1186/s12862-017-1049-1.
. PMID 28854872. doi:10.1186/s12862-017-1049-1.
- ^ 7.0 7.1 7.2 7.3 7.4 7.5 7.6 7.7 7.8 Lerosey-Aubril, Rudy; Pates, Stephen. New suspension-feeding radiodont suggests evolution of microplanktivory in Cambrian macronekton. Nature Communications. 2018-09-14, 9 (1): 3774. ISSN 2041-1723. PMC 6138677 . PMID 30218075. doi:10.1038/s41467-018-06229-7 (英語).
- ^ 8.00 8.01 8.02 8.03 8.04 8.05 8.06 8.07 8.08 8.09 8.10 8.11 8.12 8.13 8.14 8.15 8.16 8.17 8.18 8.19 8.20 8.21 8.22 8.23 8.24 8.25 8.26 8.27 8.28 8.29 8.30 Moysiuk, J.; Caron, J.-B. A new hurdiid radiodont from the Burgess Shale evinces the exploitation of Cambrian infaunal food sources. Proceedings of the Royal Society B: Biological Sciences. 2019, 286 (1908): 20191079. PMC 6710600 . PMID 31362637. doi:10.1098/rspb.2019.1079.
- ^ 9.0 9.1 9.2 9.3 Cong, Peiyun; Daley, Allison C.; Edgecombe, Gregory D.; Hou, Xianguang; Chen, Ailin. Morphology of the radiodontan Lyrarapax from the early Cambrian Chengjiang biota. Journal of Paleontology. 2016, 90 (4): 663–671. ISSN 0022-3360. doi:10.1017/jpa.2016.67 (英語).
- ^ 10.0 10.1 10.2 10.3 10.4 10.5 10.6 10.7 Daley, Allison C.; Budd, Graham E.; Caron, Jean-Bernard; Edgecombe, Gregory D.; Collins, Desmond. The Burgess Shale anomalocaridid Hurdia and its significance for early euarthropod evolution. Science. 2009, 323 (5921): 1597–1600. PMID 19299617. doi:10.1126/science.1169514.
- ^ 11.0 11.1 11.2 11.3 11.4 11.5 11.6 11.7 11.8 Cong, Peiyun; Ma, Xiaoya; Hou, Xianguang; Edgecombe, Gregory D.; Strausfeld, Nicholas J. Brain structure resolves the segmental affinity of anomalocaridid appendages. Nature. 2014, 513 (7519): 538–42. PMID 25043032. doi:10.1038/nature13486.
- ^ 12.00 12.01 12.02 12.03 12.04 12.05 12.06 12.07 12.08 12.09 12.10 12.11 12.12 12.13 12.14 12.15 12.16 12.17 12.18 12.19 12.20 12.21 12.22 12.23 Van Roy, Peter; Daley, Allison C.; Briggs, Derek E. G. Anomalocaridid trunk limb homology revealed by a giant filter-feeder with paired flaps. Nature. 2015, 522 (7554): 77–80. PMID 25762145. doi:10.1038/nature14256.
- ^ 13.0 13.1 13.2 13.3 13.4 13.5 13.6 13.7 Vinther, Jakob; Stein, Martin; Longrich, Nicholas R.; Harper, David A. T. A suspension-feeding anomalocarid from the Early Cambrian (PDF). Nature. 2014, 507 (7493): 496–499 [2020-05-30]. PMID 24670770. doi:10.1038/nature13010. (原始內容存檔 (PDF)於2021-01-22).
- ^ 14.0 14.1 Liu, Jianni; Lerosey-Aubril, Rudy; Steiner, Michael; Dunlop, Jason A.; Shu, Degan; Paterson, John R. Origin of raptorial feeding in juvenile euarthropods revealed by a Cambrian radiodontan. National Science Review. 2018-11-01, 5 (6): 863–869. ISSN 2095-5138. doi:10.1093/nsr/nwy057 (英語).
- ^ 15.0 15.1 15.2 15.3 15.4 15.5 15.6 Ortega-Hernández, Javier; Janssen, Ralf; Budd, Graham E. Origin and evolution of the panarthropod head – A palaeobiological and developmental perspective. Arthropod Structure & Development. Evolution of Segmentation. 2017-05-01, 46 (3): 354–379. ISSN 1467-8039. PMID 27989966. doi:10.1016/j.asd.2016.10.011.
- ^ 16.0 16.1 16.2 16.3 16.4 Pates, Stephen; Daley, Allison C.; Butterfield, Nicholas J. First report of paired ventral endites in a hurdiid radiodont. Zoological Letters. 2019-06-11, 5 (1): 18. ISSN 2056-306X. PMC 6560863 . PMID 31210962. doi:10.1186/s40851-019-0132-4.
- ^ 17.0 17.1 17.2 17.3 Daley, Allison C.; Budd, Graham E. New anomalocaridid appendages from the Burgess Shale, Canada. Palaeontology. 2010, 53 (4): 721–738. ISSN 1475-4983. doi:10.1111/j.1475-4983.2010.00955.x (英語).
- ^ 18.0 18.1 Daley, Allison C.; Bergström, Jan. The oral cone of Anomalocaris is not a classic peytoia. Naturwissenschaften. April 2012, 99 (6): 501–504. ISSN 0028-1042. PMID 22476406. doi:10.1007/s00114-012-0910-8 (英語).
- ^ 19.0 19.1 Strausfeld, Nicholas J.; Ma, Xiaoya; Edgecombe, Gregory D.; Fortey, Richard A.; Land, Michael F.; Liu, Yu; Cong, Peiyun; Hou, Xianguang. Arthropod eyes: The early Cambrian fossil record and divergent evolution of visual systems. Arthropod Structure & Development. August 2015, 45 (2): 152–172. PMID 26276096. doi:10.1016/j.asd.2015.07.005 (英語).
- ^ 20.0 20.1 20.2 20.3 20.4 20.5 20.6 20.7 Daley, Allison C.; Edgecombe, Gregory D. Morphology of Anomalocaris canadensis from the Burgess Shale. Journal of Paleontology. 2014, 88 (1): 68–91. doi:10.1666/13-067.
- ^ 21.0 21.1 21.2 21.3 Xian‐Guang, Hou; Bergström, Jan; Ahlberg, Per. Anomalocaris and other large animals in the lower Cambrian Chengjiang fauna of southwest China. GFF. September 1995, 117 (3): 163–183. ISSN 1103-5897. doi:10.1080/11035899509546213 (英語).
- ^ Van Roy, Peter; Daley, Allison C.; Briggs, Derek E. G. Anomalocaridids had two sets of lateral flaps. 57th Annual Meeting of The Paleontological Association. Zurich, Switzerland. 2013.
- ^ 23.0 23.1 23.2 Liu, Jianni; Lerosey-Aubril, Rudy; Steiner, Michael; Dunlop, Jason A.; Shu, Degan; Paterson, John R. Origin of raptorial feeding in juvenile euarthropods revealed by a Cambrian radiodontan. National Science Review. 2018-11-01, 5 (6): 863–869. ISSN 2095-5138. doi:10.1093/nsr/nwy057 (英語).
- ^ 24.0 24.1 24.2 Vannier, Jean; Liu, Jianni; Lerosey-Aubril, Rudy; Vinther, Jakob; Daley, Allison C. Sophisticated digestive systems in early arthropods. Nature Communications. 2014-05-02, 5 (1): 3641. ISSN 2041-1723. PMID 24785191. doi:10.1038/ncomms4641 (英語).
- ^ Daley, Allison; Antcliffe, Jonathan; Drage, Harriet; Pates, Stephen. Early fossil record of Euarthropoda and the Cambrian Explosion. Proceedings of the National Academy of Sciences. 2018-05-21, 115: 201719962 [2020-06-22]. doi:10.1073/pnas.1719962115. (原始內容存檔於2020-06-25).
- ^ 26.0 26.1 26.2 Kühl, Gabriele; Briggs, Derek E. G.; Rust, Jes. A Great-Appendage Arthropod with a Radial Mouth from the Lower Devonian Hunsrück Slate, Germany. Science. 2009-02-06, 323 (5915): 771–773. ISSN 0036-8075. PMID 19197061. doi:10.1126/science.1166586 (英語).
- ^ Van Roy, Peter; Briggs, Derek E. G. A giant Ordovician anomalocaridid. Nature. 2011-05, 473 (7348): 510–513 [2020-06-22]. ISSN 0028-0836. doi:10.1038/nature09920. (原始內容存檔於2021-02-24) (英語).
- ^ Usami, Yoshiyuki. Theoretical study on the body form and swimming pattern of Anomalocaris based on hydrodynamic simulation. Journal of Theoretical Biology. 2006-01-07, 238 (1): 11–17. ISSN 0022-5193. PMID 16002096. doi:10.1016/j.jtbi.2005.05.008.
- ^ Sheppard, K. A.; Rival, D. E.; Caron, J.-B. On the Hydrodynamics of Anomalocaris Tail Fins. Integrative and Comparative Biology. 2018-10-01, 58 (4): 703–711. ISSN 1540-7063. PMID 29697774. doi:10.1093/icb/icy014 (英語).
- ^ Daley, Allison; Drage, Harriet. The fossil record of ecdysis, and trends in the moulting behaviour of trilobites. Arthropod Structure & Development. 2015-09-01, 45 (2): 71–96. PMID 26431634. doi:10.1016/j.asd.2015.09.004.
- ^ 31.0 31.1 31.2 31.3 De Vivo, Giacinto; Lautenschlager, Stephan; Vinther, Jakob. Reconstructing anomalocaridid feeding appendage dexterity sheds light on radiodontan ecology. 2016-12-16.
- ^ Pates, S.; Daley, A. C. Caryosyntrips: a radiodontan from the Cambrian of Spain, USA and Canada. Papers in Palaeontology. 2017, 3 (3): 461–470 [2020-05-30]. ISSN 2056-2802. doi:10.1002/spp2.1084. (原始內容存檔於2019-12-24) (英語).
- ^ 33.0 33.1 33.2 33.3 33.4 33.5 33.6 33.7 33.8 Ortega-Hernández, Javier. Making sense of 'lower' and 'upper' stem-group Euarthropoda, with comments on the strict use of the name Arthropoda von Siebold, 1848. Biological Reviews of the Cambridge Philosophical Society. Dec 2014, 91 (1): 255–273. ISSN 1469-185X. PMID 25528950. doi:10.1111/brv.12168.
- ^ Ortega-Hernández, Javier. Homology of Head Sclerites in Burgess Shale Euarthropods. Current Biology. 2015-06-15, 25 (12): 1625–1631. ISSN 0960-9822. PMID 25959966. doi:10.1016/j.cub.2015.04.034.
- ^ Xian‐Guang, Hou; Bergström, Jan; Ahlberg, Per. Anomalocaris and other large animals in the lower Cambrian Chengjiang fauna of southwest China. GFF. September 1995, 117 (3): 163–183. ISSN 1103-5897. doi:10.1080/11035899509546213 (英語).
- ^ Haug, Joachim T.; Waloszek, Dieter; Maas, Andreas; Liu, Yu; Haug, Carolin. Functional morphology, ontogeny and evolution of mantis shrimp-like predators in the Cambrian: MANTIS SHRIMP-LIKE CAMBRIAN PREDATORS. Palaeontology. March 2012, 55 (2): 369–399. doi:10.1111/j.1475-4983.2011.01124.x (英語).
- ^ Smith, Martin R.; Caron, Jean-Bernard. Hallucigenia 's head and the pharyngeal armature of early ecdysozoans. Nature. June 2015, 523 (7558): 75–78. ISSN 1476-4687. PMID 26106857. doi:10.1038/nature14573 (英語).
- ^ Tanaka, Gengo; Hou, Xianguang; Ma, Xiaoya; Edgecombe, Gregory D.; Strausfeld, Nicholas J. Chelicerate neural ground pattern in a Cambrian great appendage arthropod. Nature. October 2013, 502 (7471): 364–367. ISSN 1476-4687. PMID 24132294. doi:10.1038/nature12520 (英語).
- ^ Ortega-Hernández, Javier; Lerosey-Aubril, Rudy; Pates, Stephen. Proclivity of nervous system preservation in Cambrian Burgess Shale-type deposits. Proceedings of the Royal Society B: Biological Sciences. 2019-12-18, 286 (1917): 20192370. PMC 6939931 . PMID 31822253. doi:10.1098/rspb.2019.2370.
- ^ 40.0 40.1 Wang, YuanYuan; Huang, DiYing; Hu, ShiXue. New anomalocardid frontal appendages from the Guanshan biota, eastern Yunnan. Chinese Science Bulletin. 2013-06-07, 58 (32). ISSN 1001-6538. doi:10.1007/s11434-013-5908-x.
- ^ Jiao, De-guang; Pates, Stephen; Lerosey-Aubril, Rudy; Ortega-Hernandez, Javier; Yang, Jie; Lan, Tian; Zhang, Xi-guang. The endemic radiodonts of the Cambrian Stage 4 Guanshan biota of South China. Acta Palaeontologica Polonica. 2021, 66 [2024-06-17]. ISSN 0567-7920. doi:10.4202/app.00870.2020. (原始內容存檔於2023-04-20).
- ^ Zeng, Han; Zhao, Fangchen; Zhu, Maoyan. Innovatiocaris, a complete radiodont from the early Cambrian Chengjiang Lagerstätte and its implications for the phylogeny of Radiodonta. Journal of the Geological Society. 2022-09-07. ISSN 0016-7649. doi:10.1144/jgs2021-164.
- ^ Wu, Yu; Pates, Stephen; Zhang, Mingjing; Lin, Weiliang; Ma, Jiaxin; Liu, Cong; Wu, Yuheng; Zhang, Xingliang; Fu, Dongjing. Exceptionally preserved radiodont arthropods from the lower Cambrian (Stage 3) Qingjiang Lagerstätte of Hubei, South China and the biogeographic and diversification patterns of radiodonts. Papers in Palaeontology. 2024-07, 10 (4). ISSN 2056-2799. doi:10.1002/spp2.1583.
- ^ Pates, Stephen; Lerosey-Aubril, Rudy; Daley, Allison C.; Kier, Carlo; Bonino, Enrico; Ortega-Hernández, Javier. The diverse radiodont fauna from the Marjum Formation of Utah, USA (Cambrian: Drumian). PeerJ. 2021-01-19, 9: e10509 [2021-03-19]. ISSN 2167-8359. doi:10.7717/peerj.10509. (原始內容存檔於2021-02-01) (英語).
- ^ Xian‐Guang, Hou; Bergström, Jan; Ahlberg, Per. Anomalocaris and Other Large Animals in the Lower Cambrian Chengjiang Fauna of Southwest China. GFF. 1995-09-01, 117: 163–183 [2020-06-15]. doi:10.1080/11035899509546213. (原始內容存檔於2018-07-22).
- ^ STEIN, MARTIN. A new arthropod from the Early Cambrian of North Greenland, with a ‘great appendage’-like antennula. Zoological Journal of the Linnean Society. 2010-02-26, 158 (3): 477–500 [2020-06-15]. ISSN 0024-4082. doi:10.1111/j.1096-3642.2009.00562.x. (原始內容存檔於2019-11-03) (英語).
- ^ Legg, David. Multi-Segmented Arthropods from the Middle Cambrian of British Columbia (Canada). Journal of Paleontology. 2013-05-01, 87: 493–501 [2020-06-15]. doi:10.1666/12-112.1. (原始內容存檔於2018-09-18).
- ^ C., Daley, Allison; D., Edgecombe, Gregory. Morphology of Anomalocaris canadensis from the Burgess Shale. Journal of Paleontology. [2020-06-15]. ISSN 0022-3360. (原始內容存檔於2019-10-27) (英語).
- ^ Van Roy, Peter; Daley, Allison C.; Briggs, Derek E. G. Anomalocaridid trunk limb homology revealed by a giant filter-feeder with paired flaps. Nature. 2015, 522 (7554): 77–80. ISSN 0028-0836. PMID 25762145. doi:10.1038/nature14256.
- ^ The Cambrian fossils of Chengjiang, China : the flowering of early animal life. Hou, Xianguang. Second edition. Chichester, West Sussex. ISBN 9781118896310. OCLC 970396735.
- ^ 51.0 51.1 Xianguang, Hou; Bergström, Jan; Jie, Yang. Distinguishing anomalocaridids from arthropods and priapulids. Geological Journal. 2006, 41 (3-4): 259–269 [2020-06-15]. ISSN 1099-1034. doi:10.1002/gj.1050. (原始內容存檔於2021-02-25) (德語).
- ^ Chen, Jun-yuan; Ramsköld, Lars; Zhou, Gui-qing. Evidence for Monophyly and Arthropod Affinity of Cambrian Giant Predators. Science. 1994-05-27, 264 (5163): 1304–1308 [2020-06-15]. ISSN 0036-8075. PMID 17780848. doi:10.1126/science.264.5163.1304. (原始內容存檔於2021-02-25) (英語).
- ^ Vinther, Jakob; Porras, Luis; Young, Fletcher; Budd, Graham; Edgecombe, Gregory. The mouth apparatus of the Cambrian gilled lobopodian Pambdelurion whittingtoni. Palaeontology. 2016-09-01 [2020-06-15]. doi:10.1111/pala.12256. (原始內容存檔於2018-06-12).
- ^ Zeng, Han; Zhao, Fangchen; Yin, Zongjun; Zhu, Maoyan. Morphology of diverse radiodontan head sclerites from the early Cambrian Chengjiang Lagerstätte, south-west China. Journal of Systematic Palaeontology. 2017-01-04, 16: 1 [2020-06-15]. doi:10.1080/14772019.2016.1263685. (原始內容存檔於2018-06-21).
- ^ Anomalocaridids and the origin of arthropods: the view from Chengjiang. ResearchGate. [2019-01-19]. (原始內容存檔於2019-04-17) (英語).
- ^ Guo, Jin; Pates, Stephen; Cong, Peiyun; Daley, Allison; Edgecombe, Gregory; Chen, Taimin; Hou, Xianguang. A new radiodont (stem Euarthropoda) frontal appendage with a mosaic of characters from the Cambrian (Series 2 Stage 3) Chengjiang biota. Papers in Palaeontology. 2018-08-17 [2020-06-15]. doi:10.1002/spp2.1231. (原始內容存檔於2018-09-17).
- ^ Cong, Pei-Yun; Edgecombe, Gregory D.; Daley, Allison C.; Guo, Jin; Pates, Stephen; Hou, Xian-Guang. New radiodonts with gnathobase-like structures from the Cambrian Chengjiang biota and implications for the systematics of Radiodonta. Papers in Palaeontology. 2018-06-23 [2020-06-15]. ISSN 2056-2802. doi:10.1002/spp2.1219. (原始內容存檔於2019-05-31) (英語).
- ^ Pates, S.; Botting, J. P.; McCobb, L. M. E.; Muir, L. A. A miniature Ordovician hurdiid from Wales demonstrates the adaptability of Radiodonta. Royal Society Open Science. 2020, 7 (6): 200459 [2020-06-13]. ISSN 2054-5703. doi:10.1098/rsos.200459. (原始內容存檔於2021-06-03).
- ^ Daley, Allison C.; Budd, Graham E.; Caron, Jean-Bernard; Edgecombe, Gregory D.; Collins, Desmond. The Burgess Shale Anomalocaridid Hurdia and Its Significance for Early Euarthropod Evolution. Science. 2009-03-20, 323 (5921): 1597–1600 [2020-06-22]. ISSN 0036-8075. PMID 19299617. doi:10.1126/science.1169514. (原始內容存檔於2021-02-26) (英語).
- ^ Pates, Stephen; Daley, Allison C. The Kinzers Formation (Pennsylvania, USA): the most diverse assemblage of Cambrian Stage 4 radiodonts. Geological Magazine. 2019, 156 (7): 1233–1246 [2020-07-05]. ISSN 0016-7568. doi:10.1017/S0016756818000547 (英語).
- ^ Jay,, Gould, Stephen. Wonderful life : the Burgess Shale and the nature of history First edition. New York. [2020-06-22]. ISBN 0393027058. OCLC 18983518. (原始內容存檔於2021-02-17).
- ^ Whittington, Harry Blackmore; Briggs, Derek Ernest Gilmor. The largest Cambrian animal, Anomalocaris, Burgess Shale, British-Columbia. Phil. Trans. R. Soc. Lond. B. 1985-05-14, 309 (1141): 569–609 [2020-06-22]. ISSN 0080-4622. doi:10.1098/rstb.1985.0096. (原始內容存檔於2018-10-08) (英語).
- ^ 63.0 63.1 Daley, Allison; Bergström, Jan. The oral cone of Anomalocaris is not a classic ‘‘peytoia’’. Die Naturwissenschaften. 2012-04-05, 99: 501–4 [2020-06-22]. doi:10.1007/s00114-012-0910-8. (原始內容存檔於2018-09-18).